 |
| FIG. 72– Human bone marrow. Highly magnified. |
|
| |
| Structure and Physical Properties.—Bone is one of the hardest structures of the animal body; it possesses also a certain degree of toughness and elasticity. Its color, in a fresh state, is pinkish-white externally, and deep red within. On examining a section of any bone, it is seen to be composed of two kinds of tissue, one of which is dense in texture, like ivory, and is termed compact tissue; the other consists of slender fibers and lamellæ, which join to form a reticular structure; this, from its resemblance to lattice-work, is called cancellous tissue. The compact tissue is always placed on the exterior of the bone, the cancellous in the interior. The relative quantity of these two kinds of tissue varies in different bones, and in different parts of the same bone, according as strength or lightness is requisite. Close examination of the compact tissue shows it to be extremely porous, so that the difference in structure between it and the cancellous tissue depends merely upon the different amount of solid matter, and the size and number of spaces in each; the cavities are small in the compact tissue and the solid matter between them abundant, while in the cancellous tissue the spaces are large and the solid matter is in smaller quantity. |
| Bone during life is permeated by vessels, and is enclosed, except where it is coated with articular cartilage, in a fibrous membrane, the periosteum, by means of which many of these vessels reach the hard tissue. If the periosteum be stripped from the surface of the living bone, small bleeding points are seen which mark the entrance of the periosteal vessels; and on section during life every part of the bone exudes blood from the minute vessels which ramify in it. The interior of each of the long bones of the limbs presents a cylindrical cavity filled with marrow and lined by a highly vascular areolar structure, called the medullary membrane. |
| THE STRENGTH OF BONE COMPARED WITH OTHER MATERIALS |
| Substance. |
Weight in pounds per cubic foot. |
Ultimate strength. Pounds per square inch. |
| Tension. |
Compression. |
Shear. |
| Medium steel |
490 |
65,000 |
60,000 |
40,000 |
| Granite |
170 |
1,500 |
15,000 |
2,000 |
| Oak, white |
46 |
12,500 (*12 |
7,000 |
4,000 (*13 |
| Compact bone (low) |
119 |
13,200 |
18,000 |
11,800 |
| Compact bone (high) |
…… |
17,700 |
24,000 |
7,150 |
|
| |
| Periosteum.—The periosteum adheres to the surface of each of the bones in nearly every part, but not to cartilaginous extremities. When strong tendons or ligaments are attached to a bone, the periosteum is incorporated with them. It consists of two layers closely united together, the outer one formed chiefly of connective tissue, containing occasionally a few fat cells; the inner one, of elastic fibers of the finer kind, forming dense membranous networks, which again can be separated into several layers. In young bones the periosteum is thick and very vascular, and is intimately connected at either end of the bone with the epiphysial cartilage, but less closely with the body of the bone, from which it is separated by a layer of soft tissue, containing a number of granular corpuscles or osteoblasts, by which ossification proceeds on the exterior of the young bone. Later in life the periosteum is thinner and less vascular, and the osteoblasts are converted into an epithelioid layer on the deep surface of the periosteum. The periosteum serves as a nidus for the ramification of the vessels previous to their distribution in the bone; hence the liability of bone to exfoliation or necrosis when denuded of this membrane by injury or disease. Fine nerves and lymphatics, which generally accompany the arteries, may also be demonstrated in the periosteum. |
| |
| Marrow.—The marrow not only fills up the cylindrical cavities in the bodies of the long bones, but also occupies the spaces of the cancellous tissue and extends into the larger bony canals (Haversian canals) which contain the bloodvessels. It differs in composition in different bones. In the bodies of the long bones the marrow is of a yellow color, and contains, in 100 parts, 96 of fat, 1 of areolar tissue and vessels, and 3 of fluid with extractive matter; it consists of a basis of connective tissue supporting numerous bloodvessels and cells, most of which are fat cells but some are “marrow cells,” such as occur in the red marrow to be immediately described. In the flat and short bones, in the articular ends of the long bones, in the bodies of the vertebræ, in the cranial diploë, and in the sternum and ribs the marrow is of a red color, and contains, in 100 parts, 75 of water, and 25 of solid matter consisting of cell-globulin, nucleoprotein, extractives, salts, and only a small proportion of fat. The red marrow consists of a small quantity of connective tissue, bloodvessels, and numerous cells (Fig. 72), some few of which are fat cells, but the great majority are roundish nucleated cells, the true “marrow cells” of Kölliker. These marrow cells proper, or myelocytes, resemble in appearance lymphoid corpuscles, and like them are ameboid; they generally have a hyaline protoplasm, though some show granules either oxyphil or basophil in reaction. A number of eosinophil cells are also present. Among the marrow cells may be seen smaller cells, which possess a slightly pinkish hue; these are the erythroblasts or normoblasts, from which the red corpuscles of the adult are derived, and which may be regarded as descendants of the nucleated colored corpuscles of the embryo. Giant cells (myeloplaxes, osteoclasts), large, multinucleated, protoplasmic masses, are also to be found in both sorts of adult marrow, but more particularly in red marrow. They were believed by Kölliker to be concerned in the absorption of bone matrix, and hence the name which he gave to them—osteoclasts. They excavate in the bone small shallow pits or cavities, which are named Howship’s foveolæ, and in these they are found lying. |
| |
| |
| Vessels and Nerves of Bone.—The bloodvessels of bone are very numerous. Those of the compact tissue are derived from a close and dense network of vessels ramifying in the periosteum. From this membrane vessels pass into the minute orifices in the compact tissue, and run through the canals which traverse its substance. The cancellous tissue is supplied in a similar way, but by less numerous and larger vessels, which, perforating the outer compact tissue, are distributed to the cavities of the spongy portion of the bone. In the long bones, numerous apertures may be seen at the ends near the articular surfaces; some of these give passage to the arteries of the larger set of vessels referred to; but the most numerous and largest apertures are for some of the veins of the cancellous tissue, which emerge apart from the arteries. The marrow in the body of a long bone is supplied by one large artery (or sometimes more), which enters the bone at the nutrient foramen (situated in most cases near the center of the body), and perforates obliquely the compact structure. The medullary or nutrient artery, usually accompanied by one or two veins, sends branches upward and downward, which ramify in the medullary membrane, and give twigs to the adjoining canals. The ramifications of this vessel anastomose with the arteries of the cancellous and compact tissues. In most of the flat, and in many of the short spongy bones, one or more large apertures are observed, which transmit to the central parts of the bone vessels corresponding to the nutrient arteries and veins. The veins emerge from the long bones in three places (Kölliker): (1) one or two large veins accompany the artery; (2) numerous large and small veins emerge at the articular extremities; (3) many small veins pass out of the compact substance. In the flat cranial bones the veins are large, very numerous, and run in tortuous canals in the diploic tissue, the sides of the canals being formed by thin lamellæ of bone, perforated here and there for the passage of branches from the adjacent cancelli. The same condition is also found in all cancellous tissue, the veins being enclosed and supported by osseous material, and having exceedingly thin coats. When a bone is divided, the vessels remain patulous, and do not contract in the canals in which they are contained. Lymphatic vessels, in addition to those found in the periosteum, have been traced by Cruikshank into the substance of bone, and Klein describes them as running in the Haversian canals. Nerves are distributed freely to the periosteum, and accompany the nutrient arteries into the interior of the bone. They are said by Kölliker to be most numerous in the articular extremities of the long bones, in the vertebræ, and in the larger flat bones. |
 |
| FIG. 73– Transverse section of compact tissue bone. Magnified. (Sharpey.) |
| |
| |
| Minute Anatomy.—A transverse section of dense bone may be cut with a saw and ground down until it is sufficiently thin. |
| If this be examined with a rather low power the bone will be seen to be mapped out into a number of circular districts each consisting of a central hole surrounded by a number of concentric rings. These districts are termed Haversian systems; the central hole is an Haversian canal, and the rings are layers of bony tissue arranged concentrically around the central canal, and termed lamellæ. Moreover, on closer examination it will be found that between these lamellæ, and therefore also arranged concentrically around the central canal, are a number of little dark spots, the lacunæ, and that these lacunæ are connected with each other and with the central Haversian canal by a number of fine dark lines, which radiate like the spokes of a wheel and are called canaliculi. Filling in the irregular intervals which are left between these circular systems are other lamellæ, with their lacunæ and canaliculi running in various directions, but more or less curved (Fig. 73); they are termed interstitial lamellæ. Again, other lamellæ, found on the surface of the bone, are arranged parallel to its circumference; they are termed circumferential, or by some authors primary or fundamental lamellæ, to distinguish them from those laid down around the axes of the Haversian canals, which are then termed secondary or special lamellæ. |
| The Haversian canals, seen in a transverse section of bone as round holes at or about the center of each Haversian system, may be demonstrated to be true canals if a longitudinal section be made (Fig. 74). It will then be seen that the canals run parallel with the longitudinal axis of the bone for a short distance and then branch and communicate. They vary considerably in size, some being as much as 0.12 mm. in diameter; the average size is, however, about 0.05 mm. Near the medullary cavity the canals are larger than those near the surface of the bone. Each canal contains one or two bloodvessels, with a small quantity of delicate connective tissue and some nerve filaments. In the larger ones there are also lymphatic vessels, and cells with branching processes which communicate, through the canalculi, with the branched processes of certain bone cells in the substance of the bone. Those canals near the surface of the bone open upon it by minute orifices, and those near the medullary cavity open in the same way into this space, so that the whole of the bone is permeated by a system of bloodvessels running through the bony canals in the centers of the Haversian systems. |
| The lamellæ are thin plates of bony tissue encircling the central canal, and may be compared, for the sake of illustration, to a number of sheets of paper pasted one over another around a central hollow cylinder. After macerating a piece of bone in dilute mineral acid, these lamellæ may be stripped off in a longitudinal direction as thin films. If one of these be examined with a high power of the microscope, it will be found to be composed of a finely reticular structure, made up of very slender transparent fibers, decussating obliquely; and coalescing at the points of intersection; these fibers are composed of fine fibrils identical with those of white connective tissue. The intercellular matrix between the fibers is impregnated by calcareous deposit which the acid dissolves. In many places the various lamellæ may be seen to be held together by tapering fibers, which run obliquely through them, pinning or bolting them together; they were first described by Sharpey, and were named by him perforating fibers (Fig. 75). |
 |
| FIG. 74– Section parallel to the surface from the body of the femur. X 100. a, Haversian canals; b, lacunæ seen from the side; c, others seen from the surface in lamellæ, which are cut horizontally. |
| |
 |
| FIG. 75– Perforating fibers, human parietal bone, decalcified. (H. Müller.) a, perforating fibers in situ; b, fibres drawn out of their sockets; c, sockets. |
| |
| The Lacunæ are situated between the lamellæ, and consist of a number of oblong spaces. In an ordinary microscopic section, viewed by transmitted light, they appear as fusiform opaque spots. Each lacuna is occupied during life by a branched cell, termed a bone-cell or bone-corpuscle, the processes from which extend into the canaliculi (Fig. 76). |
| The Canaliculi are exceedingly minute channels, crossing the lamellæ and connecting the lacunæ with neighboring lacunæ and also with the Haversian canal. From the Haversian canal a number of canaliculi are given off, which radiate from it, and open into the first set of lacunæ between the first and second lamellæ. From these lacunæ a second set of canaliculi is given off; these run outward to the next series of lacunæ, and so on until the periphery of the Haversian system is reached; here the canaliculi given off from the last series of lacunæ do not communicate with the lacunæ of neighboring Haversian systems, but after passing outward for a short distance form loops and return to their own lacunæ. Thus every part of an Haversian system is supplied with nutrient fluids derived from the vessels in the Haversian canal and distributed through the canaliculi and lacunæ. |
| The bone cells are contained in the lacunæ, which, however, they do not completely fill. They are flattened nucleated branched cells, homologous with those of connective tissue; the branches, especially in young bones, pass into the canaliculi from the lacunæ. |
| In thin plates of bone (as in the walls of the spaces of cancellous tissue) the Haversian canals are absent, and the canaliculi open into the spaces of the cancellous tissue (medullary spaces), which thus have the same function as the Haversian canals. |
 |
| FIG. 76– Nucleated bone cells and their processes, contained in the bone lacunæ and their canaliculi respectively. From a section through the vertebra of an adult mouse. (Klein and Noble Smith.) |
| |
| |
| Chemical Composition.—Bone consists of an animal and an earthy part intimately combined together. |
| The animal part may be obtained by immersing a bone for a considerable time in dilute mineral acid, after which process the bone comes out exactly the same shape as before, but perfectly flexible, so that a long bone (one of the ribs, for example) can easily be tied in a knot. If now a transverse section is made (Fig. 77) the same general arrangement of the Haversian canals, lamellæ, lacunæ, and canaliculi is seen. |
| The earthy part may be separately obtained by calcination, by which the animal matter is completely burnt out. The bone will still retain its original form, but it will be white and brittle, will have lost about one-third of its original weight, and will crumble down with the slightest force. The earthy matter is composed chiefly of calcium phosphate, about 58 per cent. of the weight of the bone, calcium carbonate about 7 per cent., calcium fluoride and magnesium phosphate from 1 to 2 per cent. each and sodium chloride less than 1 per cent.; they confer on bone its hardness and rigidity, while the animal matter (ossein) determines its tenacity. |
| |
| Ossification.—Some bones are preceded by membrane, such as those forming the roof and sides of the skull; others, such as the bones of the limbs, are preceded by rods of cartilage. Hence two kinds of ossification are described: the intramembranous and the intracartilaginous. |
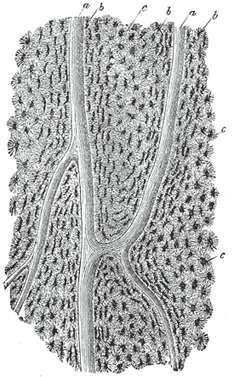
| INTRAMEMBRANOUS OSSIFICATION.—In the case of bones which are developed in membrane, no cartilaginous mould precedes the appearance of the bony tissue. The membrane which occupies the place of the future bone is of the nature of connective tissue, and ultimately forms the periosteum; it is composed of fibers and granular cells in a matrix. The peripheral portion is more fibrous, while, in the interior the cells or osteoblasts predominate; the whole tissue is richly supplied with blood vessels. At the outset of the process of bone formation a little network of spicules is noticed radiating from the point or center of ossification. These rays consist at their growing points of a network of fine clear fibers and granular corpuscles with an intervening ground substance (Fig. 78). The fibers are termed osteogenetic fibers, and are made up of fine fibrils differing little from those of white fibrous tissue. The membrane soon assumes a dark and granular appearance from the deposition of calcareous granules in the fibers and in the intervening matrix, and in the calcified material some of the granular corpuscles or osteoblasts are enclosed. By the fusion of the calcareous granules the tissue again assumes a more transparent appearance, but the fibers are no longer so distinctly seen. The involved osteoblasts from the corpuscles of the future bone, the spaces in which they are enclosed constituting the lacunæ. As the osteogenetic fibers grow out to the periphery they continue to calcify, and give rise to fresh bone spicules. Thus a network of bone is formed, the meshes of which contain the bloodvessels and a delicate connective tissue crowded with osteoblasts. The bony trabeculæ thicken by the addition of fresh layers of bone formed by the osteoblasts on their surface, and the meshes are correspondingly encroached upon. Subsequently successive layers of bony tissue are deposited under the periosteum and around the larger vascular channels which become the Haversian canals, so that the bone increases much in thickness. |
 |
| FIG. 77– Transverse section of body of human fibula, decalcified. X 250. |
| |
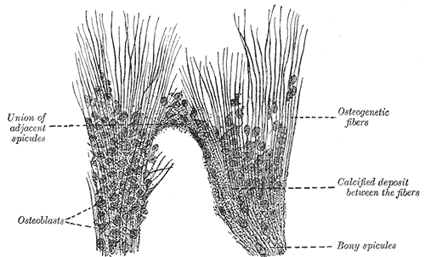 |
| FIG. 78– Part of the growing edge of the developing parietal bone of a fetal cat. (After J. Lawrence.) |
| |
| INTERCARTILAGINOUS OSSIFICATION.—Just before ossification begins the mass is entirely cartilaginous, and in a long bone, which may be taken as an example, the process commences in the center and proceeds toward the extremities, which for some time remain cartilaginous. Subsequently a similar process commences in one or more places in those extremities and gradually extends through them. The extremities do not, however, become joined to the body of the bone by bony tissue until growth has ceased; between the body and either extremity a layer of cartilaginous tissue termed the epiphysial cartilage persists for a definite period. |
| The first step in the ossification of the cartilage is that the cartilage cells, at the point where ossification is commencing and which is termed a center of ossification, enlarge and arrange themselves in rows (Fig. 79). The matrix in which they are imbedded increases in quantity, so that the cells become further separated from each other. A deposit of calcareous material now takes place in this matrix, between the rows of cells, so that they become separated from each other by longitudinal columns of calcified matrix, presenting a granular and opaque appearance. Here and there the matrix between two cells of the same row also becomes calcified, and transverse bars of calcified substance stretch across from one calcareous column to another. Thus there are longitudinal groups of the cartilage cells enclosed in oblong cavities, the walls of which are formed of calcified matrix which cuts off all nutrition from the cells; the cells, in consequence, atrophy, leaving spaces called the primary areolæ. |
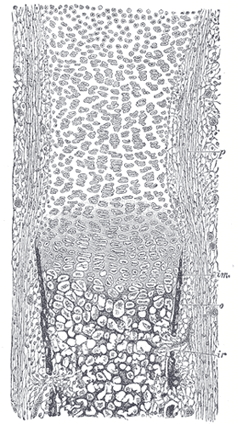 |
| FIG. 79– Section of fetal bone of cat. ir. Irruption of the subperiosteal tissue. p. Fibrous layer of the periosteum. o. Layer of osteoblasts. im. Subperiosteal bony deposit. (From Quain’s “Anatomy,” E. A. Schäfer.) |
| |
| At the same time that this process is going on in the center of the solid bar of cartilage, certain changes are taking place on its surface. This is covered by a very vascular membrane, the perichondrium, entirely similar to the embryonic connective tissue already described as constituting the basis of membrane bone; on the inner surface of this—that is to say, on the surface in contact with the cartilage—are gathered the formative cells, the osteoblasts. By the agency of these cells a thin layer of bony tissue is formed between the perichondrium and the cartilage, by the intramembranous mode of ossification just described. There are then, in this first stage of ossification, two processes going on simultaneously: in the center of the cartilage the formation of a number of oblong spaces, formed of calcified matrix and containing the withered cartilage cells, and on the surface of the cartilage the formation of a layer of true membrane bone. The second stage consists in the prolongation into the cartilage of processes of the deeper or osteogenetic layer of the perichondrium, which has now become periosteum (Fig. 79, ir). The processes consist of bloodvessels and cells—osteoblasts, or bone-formers, and osteoclasts, or bone-destroyers. The latter are similar to the giant cells (myeloplaxes) found in marrow, and they excavate passages through the new-formed bony layer by absorption, and pass through it into the calcified matrix (Fig. 80). Wherever these processes come in contact with the calcified walls of the primary areolæ they absorb them, and thus cause a fusion of the original cavities and the formation of larger spaces, which are termed the secondary areolæ or medullary spaces. These secondary spaces become filled with embryonic marrow, consisting of osteoblasts and vessels, derived, in the manner described above, from the osteogenetic layer of the periosteum (Fig. 80). |
| Thus far there has been traced the formation of enlarged spaces (secondary areolæ), the perforated walls of which are still formed by calcified cartilage matrix, containing an embryonic marrow derived from the processes sent in from the osteogenetic layer of the periosteum, and consisting of bloodvessels and osteoblasts. The walls of these secondary areolæ are at this time of only inconsiderable thickness, but they become thickened by the deposition of layers of true bone on their surface. This process takes place in the following manner: Some of the osteoblasts of the embryonic marrow, after undergoing rapid division, arrange themselves as an epithelioid layer on the surface of the wall of the space (Fig. 81). This layer of osteoblasts forms a bony stratum, and thus the wall of the space becomes gradually covered with a layer of true osseous substance in which some of the bone-forming cells are included as bone corpuscles. The next stage in the process consists in the removal of these primary bone spicules by the osteoclasts. One of these giant cells may be found lying in a Howship’s foveola at the free end of each spicule. The removal of the primary spicules goes on pari passu with the formation of permanent bone by the periosteum, and in this way the medullary cavity of the body of the bone is formed. |
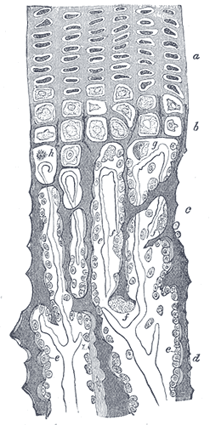 |
| FIG. 80– Part of a longitudinal section of the developing femur of a rabbit. a. Flattened cartilage cells. b. Enlarged cartilage cells. c, d. Newly formed bone. e. Osteoblasts. f. Giant cells or osteoclasts. g, h. Shrunken cartilage cells. (From “Atlas of Histology,” Klein and Noble Smith.) |
| |
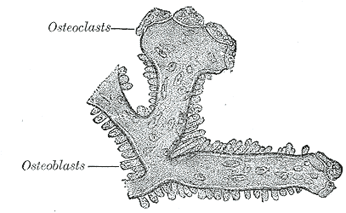 |
| FIG. 81– Osteoblasts and osteoclasts on trabecula of lower jaw of calf embryo. (Kölliker.) |
| |
| This series of changes has been gradually proceeding toward the end of the body of the bone, so that in the ossifying bone all the changes described above may be seen in different parts, from the true bone at the center of the body to the hyaline cartilage at the extremities. |
| While the ossification of the cartilaginous body is extending toward the articular ends, the cartilage immediately in advance of the osseous tissue continues to grow until the length of the adult bone is reached. |
| During the period of growth the articular end, or epiphysis, remains for some time entirely cartilaginous, then a bony center appears, and initiates in it the process of intracartilaginous ossification; but this process never extends to any great distance. The epiphysis remains separated from the body by a narrow cartilaginous layer for a definite time. This layer ultimately ossifies, the distinction between body and epiphysis is obliterated, and the bone assumes its completed form and shape. The same remarks also apply to such processes of bone as are separately ossified, e.g., the trochanters of the femur. The bones therefore continue to grow until the body has acquired its full stature. They increase in length by ossification continuing to extend behind the epiphysial cartilage, which goes on growing in advance of the ossifying process. They increase in circumference by deposition of new bone, from the deeper layer of the periosteum, on their external surface, and at the same time an absorption takes place from within, by which the medullary cavities are increased. |
| The permanent bone formed by the periosteum when first laid down is cancellous in structure. Later the osteoblasts contained in its spaces become arranged in the concentric layers characteristic of the Haversian systems, and are included as bone corpuscles. |
| The number of ossific centers varies in different bones. In most of the short bones ossification commences at a single point near the center, and proceeds toward the surface. In the long bones there is a central point of ossification for the body or diaphysis: and one or more for each extremity, the epiphysis. That for the body is the first to appear. The times of union of the epiphyses with the body vary inversely with the dates at which their ossifications began (with the exception of the fibula) and regulate the direction of the nutrient arteries of the bones. Thus, the nutrient arteries of the bones of the arm and forearm are directed toward the elbow, since the epiphyses at this joint become united to the bodies before those at the opposite extremities. In the lower limb, on the other hand, the nutrient arteries are directed away from the knee: that is, upward in the femur, downward in the tibia and fibula; and in them it is observed that the upper epiphysis of the femur, and the lower epiphyses of the tibia and fibula, unite first with the bodies. Where there is only one epiphysis, the nutrient artery is directed toward the other end of the bone; as toward the acromial end of the clavicle, toward the distal ends of the metacarpal bone of the thumb and the metatarsal bone of the great toe, and toward the proximal ends of the other metacarpal and metatarsal bones. |
| Parsons (*14 groups epiphyses under three headings, viz.: (1) pressure epiphyses, appearing at the articular ends of the bones and transmitting “the weight of the body from bone to bone;” (2) traction epiphyses, associated with the insertion of muscles and “originally sesamoid structures though not necessarily sesamoid bones;” and (3) atavistic epiphyses, representing parts of the skeleton, which at one time formed separate bones, but which have lost their function, “and only appear as separate ossifications in early life.” |
