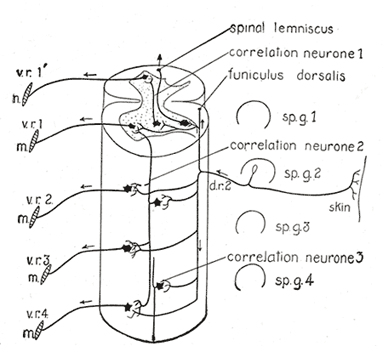
 |
| FIG. 758– Diagram of the spinal cord reflex apparatus. Some of the connections of a single afferent neuron from the skin (d.r.2) are indicated: d.r.2, dorsal root from second spinal ganglion; m, muscles; sp.g.1 to sp.g.4, spinal ganglia; v.r.1' to v.r.4, ventral roots. (After Herrick.) |
|
| The typical spinal nerve consists of at least four types of fibers, the somatic sensory, sympathetic afferent or sensory, somatic motor and sympathetic efferent or preganglionic. The somatic sensory fibers, afferent fibers, arise from cells in the spinal ganglia and are found in all the spinal nerves, except occasionally the first cervical, and conduct impulses of pain, touch and temperature from the surface of the body through the posterior roots to the spinal cord and impulses of muscle sense, tendon sense and joint sense from the deeper structures. The sympathetic afferent fibers, conduct sensory impulses from the viscera through the rami communicantes and posterior roots to the spinal cord. They are probably limited to the white rami connected with the spinal nerves in two groups, viz., the first thoracic to the second lumbar and the second sacral to the fourth sacral nerves. The somatic motor fibers, efferent fibers, arise from cells in the anterior column of the spinal cord and pass out through the anterior roots to the voluntary muscles. The sympathetic efferent fibers, probably arise from cells in the lateral column or the base of the anterior column and emerge through the anterior roots and white rami communicantes. These are preganglionic fibers which end in various sympathetic ganglia from which postganglionic fibers conduct the motor impulses to the smooth muscles of the viscera and vessels and secretory impulses to the glands. These fibers are also limited to two regions, the first thoracic to the second lumbar and the second sacral to the fourth sacral nerves. |
| The afferent fibers which pass into the spinal cord establish various types of connections, some within the cord itself for spinal reflexes, others for reflexes connected with higher centers in the brain, while still others conduct impulses of conscious sensation by a series of neurons to the cerebral cortex. |
| |
| |
| The Intrinsic Spinal Reflex Paths.—The collaterals and terminals of the ascending and descending branches of the posterior root fibers which leave the fasciculus cuneatus to enter the gray matter of the spinal cord end in various ways. Many end in the dorsal column, some near its apex, others in the substance of Rolando, others in the intermediate region between the dorsal and ventral columns, others traverse the whole thickness of the gray matter to reach the ventral column, others end in the dorsal nucleus, and others pass through the gray commissure to the dorsal column of the opposite side. All of these collaterals and terminals end in connection with cells or dendrites of cells in the gray columns. The axons of these cells have various destinations, some pass out into the lateral and ventral funiculi and turn upward to reach the brain. Those concerned with the intrinsic spinal reflexes come into relation either directly or indirectly with motor cells in the anterior column. It is very unlikely that either the terminals or collaterals of the dorsal root fibers effect simple direct connections with the motor cells of the ventral column, there is at least one if not several intercalated neurons in the path. These intercalated or correlation neurons may have short axons that do not pass out of the gray matter or the axons may pass out into the proper fasciculi and extend for varying distances up and down or in both directions giving off collaterals and finally terminating in the gray matter of the same or the opposite side. The shortest fibers of the proper fasciculi lie close to the gray matter, the longest ones are nearer the periphery of the proper fasciculi and are more or less intermingled with the long ascending and descending fasciculi which occupy the more marginal regions of the spinal cord. |
| Each sensory neuron, with its ascending and descending branches, giving off as it does many collaterals into the gray matter, each one of which may form a synapse with one or several correlation neurons, is thus brought into relation with many correlation neurons and each one of these in turn, with its ascending and descending branches and their numerous collaterals, is brought into relation, either directly or through the intercalation of additional correlation neurons, with great numbers of motor cells in the anterior column. The great complexity of these so-called simple reflex mechanisms, in the least complex portion of the nervous system the spinal cord, renders them extremely difficult of exact analysis. |
| The association or correlation neurons are concerned not only with the reflex mechanisms of the spinal cord but play an equally important role in the transmission of impulses from the higher centers in the brain to the motor neurons of the spinal cord. |
| The complex mechanisms just described are probably concerned not so much in the contraction of individual muscles as in the complicated action of groups of muscles concerned in the enormous number of movements, which the limbs and trunk exhibit in the course of our daily life. |
| |
| Sensory Pathways from the Spinal Cord to the Brain.—The posterior root fibers conducting the impulses of conscious muscle sense, tendon sense and joint sense, those impulses which have to do with the coördination and adjustment of muscular movements, ascend in the fasciculus gracilis and fasciculus cuneatus to the nucleus gracilis and nucleus cuneatus in the medulla oblongata (Fig. 759). |
| In the nucleus gracilis and nucleus cuneatus synaptic relations are found with neurons whose cell bodies are located in these nuclei and whose axons pass by way of the internal arcuate fibers, cross in the raphé to the opposite side in the region between the olives and turn abruptly upward to form the medial lemniscus or medial fillet. The medial fillet passes upward in the ventral part of the formatio reticularis through the medulla oblongata, pons and mid-brain to the principal sensory nucleus of the ventro-lateral region of the thalamus. Here the terminals form synapses with neurons of the third order whose axons pass through the internal capsule and corona radiata to the somatic sensory area of the cortex in the post-central gyrus. |
| Fibers conducting the impulses of unconscious muscle sense pass to the cerebellum partly by way of the fasciculus gracilis and fasciculus cuneatus to the nucleus gracilis and nucleus cuneatus, thence neurons of the second order convey the impulses either via the dorsal external arcuate fibers directly into the inferior peduncle of the cerebellum or via the ventral external arcuate fibers which are continued from the internal arcuate fibers through the ventral part of the raphé and after crossing the midline emerge on the surface of the medulla in the ventral sulcus between the pyramids or in the groove between the pyramid and the olive. They pass over the lateral surface of the medulla and olive to reach the inferior peduncle through which they pass to the cerebellum. |
| Other fibers conducting impulses of unconscious muscle sense pass upward in the dorsal spinocerebellar fasciculus, which arises from cells in the nucleus dorsalis. The posterior root fibers conducting these impulses pass into the fasciculus cuneatus and the collaterals from them to the nucleus dorsalis are said to come almost exclusively from the middle area of the fasciculus cuneatus. They form by their multiple division baskets about the individual cells of the nucleus dorsalis, each fiber coming in relation with the bodies and dendrites of several cells. The axons of the second order pass into the dorsal spinocerebellar fasciculus of the same side and ascend along the lateral surface of the spinal cord and medulla oblongata until they arrive at the level of the olive, they then curve backward beneath the external arcuate fibers into the inferior peduncle and pass into the cerebellum. Here they give off collaterals to the dentate nucleus and finally terminate in the cortex of the dorsal and superior portion of the vermis, partly on the same side, but to a great extent by way of a large commissure to the opposite side. The fibers lose their myelin sheaths as they enter the gray substance and terminate by end ramifications among the nerve cells and their processes. Some of the fibers are said to end in the nucleus dentatus and the roof nuclei of the cerebellum (the nucleus globosus, nucleus emboliformis and nucleus fastigius) and others pass through them to terminate in the inferior vermis. A few fibers of the dorsal spinocerebellar fasciculus are said not to enter the inferior peduncle but to pass with the ventral spinocerebellar fasciculus. The cerebellar reflex are is supposed to be completed by the fibers of the superior peduncle which pass from the cerebellum to the red nucleus of the mid-brain where some of their terminals and collaterals form synapses with neurons whose axons descend to the spinal cord in the rubrospinal fasciculus. The terminal and collaterals of this fasciculus end either directly or indirectly about the motor cells in the anterior column. |
 |
| FIG. 759– The sensory tract. (Modified from Poirier.) |
| |
| The ventral spinocerebellar fasciculus, since most of its fibers pass to the cerebellum, is also supposed to be concerned in the conduction of unconscious muscle sense. The location of its cells of origin is uncertain. They are probably in or near the dorsal nucleus of the same and the opposite side; various other locations are given, the dorsal column, the intermediate zone of the gray matter and the central portion of the anterior column. The neurons of the first order whose central fibers enter the fasciculus cuneatus from the dorsal roots send collaterals and terminals to form synapses with these cells. The fibers which come from the opposite gray columns cross some in the white and some in the gray commissure and pass with fibers from the same side through the lateral funiculus to the marginal region ventral to the dorsal spinocerebellar fasciculus. The fasciculus begins about the level of the third lumbar nerve and continues upward on the lateral surface of the spinal cord and medulla oblongata until it passes under cover of the external arcuate fibers. It passes just dorsal to the olive and above this joins the lateral edge of the lateral lemniscus along which it runs, ventral to the roots of the trigeminal nerve, almost to the level of the superior colliculus, it then crosses over the superior peduncle, turns abruptly backward along its medial border, enters the cerebellum with it and ends in the vermis of the same and the opposite side. Some of its fibers are said to join the dorsal spinocerebellar fasciculus in the medulla oblongata and enter the cerebellum through the inferior peduncle. A number of fibers are said to continue upward in the dorsolateral part of the tegmentum as far as the superior colliculus and a few pass to the thalamus. They probably form part of the sensory or higher reflex path. |
| The posterior root fibers conducting impulses of pain and temperature probably terminate in the posterior column or the intermediate region of the gray matter soon after they enter the spinal cord. The neurons of the second order are supposed to pass through the anterior commissure to the superficial antero-lateral fasciculus (tract of Gowers) and pass upward in that portion of it known as the lateral spinothalamic fasciculus. This fasciculus lies along the medial side of the ventral spinocerebellar fasciculus. It is stated by some authors that the pain fibers pass upward in the antero-lateral ground bundles. In some of the lower mammals this pathway carries the pain fibers upward by a series of neurons some of which cross to the opposite side, so that in part there is a double path. In man, however, the lateral spinothalamic fasciculus is probably the most important pathway. On reaching the medulla these fibers continue upward through the formatio reticularis in the neighborhood of the median fillet to the thalamus, probably its ventro-lateral region. Whether higher neurons convey the pain impulses to the cortex through the internal capsule is uncertain. The pathway is probably more complex and Head is of the opinion that our sensations of pain are essentially thalamic. The pain and temperature pathways in the lateral spinothalamic fasciculus are not so closely intermingled but that one can be destroyed without injury to the other. |
| Ransom suggests that the non-medullated fibers of the posterior roots, which turn into Lissauer’s tract and ascend or descend for short distances not exceeding one or two segments and finally end in the substantia gelatinosa, are in part at least pain fibers and that the fasciculus of Lissauer and the substantia gelatinosa represent part of the mechanism for reflexes associated with pain conduction and reception while the fibers to the higher centers pass up in the spinothalamic tract. |
| The fibers of tactile discrimination, according to Head and Thompson, pass up in the fasciculus cuneatus and fasciculus gracilis of the same side and follow the path of the muscle-sense fibers. The axons of the second order arising in the nucleus cuneatus and gracilis cross with the internal arcuate fibers and ascend to the thalamus with the medial lemniscus, thence by neurons of higher order the impulses are carried to the somatic sensory area of the cortex through the internal capsule. The other touch fibers, shortly after entering the spinal cord, terminate in the dorsal column or intermediate gray matter. Neurons of the second order send their axons through the anterior commissure to pass upward in the antero-lateral funiculus probably in the ventral spinothalamic fasciculus. In the medulla they join or pass upward in the neighborhood of the medial lemniscus to the thalamus and thence by neurons of higher order to the somatic sensory area of the cortex. |
| The remaining ascending fasciculi form a part of the complex known as the superficial antero-lateral fasciculus (tract of Gowers). The spinotectal fasciculus, as its name indicates, is supposed to have its origin in the gray matter of the cord and terminations in the superior and inferior (?) colliculi of the mid-brain serving for reflexes between the cord and the visceral and auditory centers of the mid-brain. |
| The spino-olivary fasciculus (olivospinal; bulbospinal, Helweg’s bundle) is likewise of unknown constitution and function; there is uncertainty even in regard to the direction of its fibers. |
| Sympathetic afferent fibers (visceral afferent; viscero-sensory; splanchnic afferent) enter the spinal cord by the posterior roots of the thoracic and first two or three lumbar nerves and the second to the fourth sacral nerves. The fibers pass to these nerves from the peripheral sympathetic system through the white rami communicantes. Some of the cell bodies of these afferent fibers are located in the spinal ganglia and others are in the sympathetic ganglia. Some of the afferent sympathetic fibers end about the cell bodies of somatic sensory neurons and visceral impulses are thus transmitted to these neurons which conduct them as well as their own special impulses to the spinal cord. Other sympathetic afferent neurons whose cell bodies are located in the spinal ganglia send collaterals to neighboring cells of somatic sensory neurons and thus have a double path of transmission to the spinal cord. Such an arrangement provides a mechanism for some of the referred pains. |
| These sympathetic afferent fibers presumably divide on entering the spinal cord into ascending and descending branches. Their distribution and termination within the spinal cord are unknown. Some of them probably eventually come into relation with the sympathetic efferent fibers whose cell bodies are located in the lateral column. Our knowledge concerning both the termination and origin of these fibers is very unsatisfactory. |
| The sympathetic efferent fibers (splanchnic motor; viscero-motor; preganglionic fibers) are supposed to arise from cells in the intermediate zone between the dorsal and ventral gray columns and in the intermedio-lateral column at the margin of the lateral column. These preganglionic sympathetic fibers are not distributed throughout the entire series of spinal nerves but are confined to two groups, the thoraco-lumbar from the first thoracic to the second or third lumbar nerves and the sacral group from the second to the fourth sacral nerves. They pass out with the anterior root fibers and through the rami communicantes to end in sympathetic ganglia. The impulses are distributed from cells in these ganglia through postganglionic fibers to the smooth muscles and glands. The thoraco-lumbar outflow and the sacral outflow form two distinct functional groups which are considered more fully under the sympathetic system. |
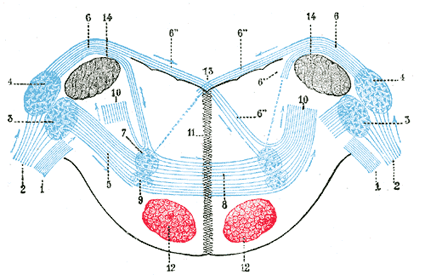 |
| FIG. 760– Terminal nuclei of the cochlear nerve, with their upper connections. (Schematic.) The vestibular nerve with its terminal nuclei and their efferent fibers have been suppressed. On the other hand, in order not to obscure the trapezoid body, the efferent fibers of the terminal nuclei on the right side have been resected in a considerable portion of their extent. The trapezoid body, therefore, shows only one-half of its fibers, viz., those which come from the left. 1. Vestibular nerve, divided at its entrance into the medulla oblongata. 2. Cochlear nerve. 3. Accessory nucleus of acoustic nerve. 4. Tuberculum acusticum. 5. Efferent fibers of accessory nucleus. 6. Efferent fibers of tuberculum acusticum, forming the striae medullares, with 6’, their direct bundle going to the superior olivary nucleus of the same side; 6’’, their decussating bundles going to the superior olivary nucleus of the opposite side. 7. Superior olivary nucleus. 8. Trapezoid body. 9. Trapezoid nucleus. 10. Central acoustic tract (lateral lemniscus). 11. Raphé. 12. Cerebrospinal fasciculus. 13. Fourth ventricle. 14. Inferior peduncle. (Testut.) |
|
| The cranial nerves are more varied in their composition than the spinal nerves. Some, for example, contain somatic motor fibers only, others contain the various types of fibers found in the spinal nerves, namely, somatic motor, sympathetic efferent, somatic sensory and sympathetic sensory. In addition there are included the nerves of the special senses, namely, the nerves of smell, sight, hearing, equilibration and taste. |
| The Hypoglossal Nerve (XII cranial) consists of somatic motor fibers only and supplies the muscles of the tongue. Its axons arise from cells in the hypoglossal nucleus and pass forward between the white reticular formation and the gray reticular formation to emerge from the antero-lateral sulcus of the medulla. The hypoglossal nuclei of the two sides are connected by many commissural fibers and also by dendrites of motor cells which extend across the midline to the opposite nucleus. The hypoglossal nucleus receives either directly or indirectly numerous collaterals and terminals from the opposite pyramidal tract (cortico-bulbar or cerebrobulbar fibers) which convey voluntary motor impulses from the cerebral cortex. Many reflex collaterals enter the nucleus from the secondary sensory paths of the trigeminal and vagus and probably also from the nervus intermedius and the glossopharyngeal. Collaterals from the posterior longitudinal bundle and the ventral longitudinal bundle are said to pass to the nucleus. |
| The Accessory Nerve (XI cranial) contains somatic motor fibers. The spinal part arises from lateral cell groups in the anterior column near its dorso-lateral margin in the upper five or six segments of the cord, its roots pass through the lateral funiculus to the lateral surface of the cord. It supplies the Trapezius and Sternocleidomastoideus. The cranial part arises from the nucleus ambiguus, the continuation in the medulla oblongata of the lateral cell groups of the anterior column of the spinal cord from which the spinal part has origin. The upper part of the nucleus ambiguus gives motor fibers to the vagus and glossopharyngeal nerves. The cranial part sends it fibers through the vagus to the laryngeal nerves to supply the muscles of the larynx. The root fibers of the cranial part of the accessory nerve pass anterior to the spinal tract of the trigeminal while those of the vagus pass through or dorsal to the trigeminal root, and emerge in the line of the postero-lateral sulcus. The nucleus of origin of the spinal part undoubtedly receives either directly or indirectly terminals and collaterals controlling voluntary movements from the pyramidal tracts. It is probable that terminals and collaterals reach the nucleus either directly or indirectly from the rubrospinal and the vestibulospinal tracts. It is also connected indirectly with the spinal somatic sensory nerves by association fibers of the proper fasciculi. The cranial part receives indirectly or directly terminals and collaterals from the opposite pyramidal tract and form the terminal sensory nuclei of the cranial nerves. A few fibers of the cranial part are said to arise in the dorsal nucleus of the vagus and are thus sympathetic efferent. They are said to join the vagus nerve. |
| The Vagus Nerve (X cranial) contains somatic sensory, sympathetic afferent, somatic motor, sympathetic efferent and (taste fibers?). The afferent fibers (somatic sensory, sympathetic, and taste) have their cells of origin in the jugular ganglion and in the nodosal ganglion (ganglion of the trunk) and on entering the medulla divide into ascending and descending branches as do the sensory fibers of the posterior roots of the spinal nerves after they enter the spinal cord. |
| (1) The somatic sensory fibers are few in number, convey impulses from a limited area of the skin on the back of the ear and posterior part of the external auditory meatus, and probably join the spinal tract of the trigeminal nerve to terminate in its nucleus. Connections are probably established through the central path of the trigeminal with the thalamus and somatic sensory area of the cortex for the conscious recognition of impulses. The descending fibers in the spinal tract of the trigeminal terminating in the nucleus of the tract probably establish relations through connecting neurons with motor nuclei in the anterior column of the spinal cord and with motor nuclei of the medulla. |
| (2) The sympathetic afferent fibers are usually described as terminating in the dorsal nucleus of the vagus and glossopharyngeal. Some authors, however, believe they join the tractus solitarius and terminate in its nucleus. These afferent fibers convey impulses from the heart, the pancreas, and probably from the stomach, esophagus and respiratory tract. Their terminals in the dorsal nucleus come into relation with neurons whose axons probably descend into the spinal cord, conveying impulses to the motor nuclei supplying fibers to the muscles of respiration, i. e., the phrenic nerve and the nerves to the intercostal and levatores costarum muscles. Other axons probably convey vasomotor impulses to certain sympathetic efferent neurons throughout the spinal cord. The dorsal nucleus (nucleus of the ala cinerea) and the posterior continuation of it into the commissural nucleus of the ala cinerea constitute probably the so-called respiratory and vaso-motor center of the medulla. The shorter reflex neurons of the dorsal nucleus probably effect connections either directly or indirectly with motor cells of the vagus itself and other cranial nerves. |
| (3) Taste fibers conducting impulses from the epiglottis and larynx are supposed to pass in the vagus and to join the tractus solitarius, finally terminating in the nucleus of the tractus solitarius. It is not certain that this nucleus represents the primary terminal center for taste and some authors maintain that the taste fibers terminate in the dorsal nucleus. The secondary ascending pathways from the primary gustatory nucleus to the cortex as well as the location of the cortical center for taste are unknown. A gustatory center has been described near the anterior end of the temporal lobe. The nucleus of the tractus solitarius is connected with motor centers of the pons, medulla and spinal cord for the reactions of mastication and swallowing. |
| (4) Somatic motor fibers to the cross striated muscles of the pharynx and larynx arise in the nucleus ambiguus. This nucleus undoubtedly receives either directly or indirectly collaterals or terminals from the opposite pyramidal tract controlling the voluntary movements of the pharynx and larynx. The reflex pathways conveying impulses from the terminal sensory nuclei are unknown, but probably form part of the intricate maze of fibers constituting the reticular formation. |
| (5) Sympathetic efferent fibers arise from cells in the dorsal nucleus (nucleus of the ala cinerea). These are preganglionic fibers of the sympathetic system and all terminate in sympathetic ganglia from which postganglionic fibers are distributed to various organs, i. e., motor fibers to the esophagus, stomach, small intestine, gallbladder, and to the lungs; inhibitory fibers to the heart; secretory fibers to the stomach and pancreas. The dorsal nucleus not only receives terminals of sympathetic afferent fibers for reflexes but undoubtedly receives terminals and collaterals from many other sources, but the exact pathways are at present unknown. |
| The Glossopharyngeal Nerve (IX cranial) is similar to the vagus nerve as regards its central connections and is usually described with it. It contains somatic sensory, sympathetic afferent, taste, somatic motor and sympathetic efferent fibers. The afferent sensory fibers arise from cells in the superior ganglion and in the petrosal ganglion. The same uncertainty exists concerning the nuclei of termination and nuclei of origin of the various components as for the vagus. |
| (1) The somatic sensory fibers are few in number. Some are distributed with the auricular branch of the vagus to the external ear; others probably pass to the pharynx and fauces. They are supposed to join the spinal tract of the trigeminal and terminate in its nucleus. The connections are similar to those of the somatic sensory fibers of the vagus. |
| (2) Sympathetic afferent fibers from the pharynx and middle ear are supposed to terminate in the dorsal nucleus. Connections are probably established with motor nuclei concerned in chewing and swallowing; very little is known, however, about the connections with other parts of the brain. |
| (3) Taste fibers from the tongue probably terminate in the nucleus of the tractus solitarius. These fibers together with similar fibers from the facial (nervus intermedius) and the vagus are supposed to form the tractus solitarius and terminate in its nucleus. The central connections have been considered under the vagus. |
| (4) Somatic motor fibers to the Sylopharyngeus muscle arise in the upper end of the nucleus ambiguus. The existence of these fibers in the roots of the glossopharyngeal is uncertain, as there are other paths by which such fibers might reach the glossopharyngeal from the vagus. The sources of impulses passing to the nucleus ambiguus are considered under the vagus. |
| (5) Sympathetic efferent fibers (motor and secretory fibers) arise from the nucleus dorsalis. Some authors believe that the secretory fibers to the parotid gland arise from a distinct nucleus, the inferior salivatory nucleus, situated near the dorsal nucleus. The preganglionic fibers from this nucleus terminate in the otic ganglion; the postganglionic fibers from the otic ganglion pass to the parotid gland. |
| The Acoustic Nerve (VIII cranial) consists of two distinct nerves the cochlear nerve, the nerve of hearing, and the vestibular nerve, the nerve of equilibration. |
| |
| The Cochlear Nerve arises from bipolar cells in the spiral ganglion of the cochlea; the peripheral fibers end in the organ of Corti, the central fibers bifurcate as they enter the cochlear nucleus; the short ascending branches end in the ventral portion of the nucleus, the longer descending branches terminate in the dorsal portion of the nucleus. From the dorsal portion of the cochlear nucleus axons arise which pass across the dorsal aspect of the inferior peduncle and the floor of the fourth ventricle, the striæ medullares, to the median sulcus. Here they dip into the substance of the pons, cross the median plane, and join the lateral lemniscus. Some of the fibers terminate in the superior olivary nucleus. The fibers of the striæ medullares are not always visible on the floor of the rhomboid fossa. From the ventral portion of the cochlear nucleus axons pass into the trapezoid body, here some of them end in the superior olivary nucleus of the same side, others cross the midline and end in the superior olivary nucleus of the opposite side or pass by these nuclei, giving off collaterals to them, and join the lateral lemniscus. Other fibers either terminate in or give off collaterals to the nucleus of the trapezoid body of the same or the opposite side. Other fibers from the ventral portion of the cochlear nucleus pass dorsal to the inferior peduncle and then dip into the substance of the pons to join the trapezoid body or the superior olivary nucleus of the same side. From the superior olivary nucleus of the same and opposite sides axons join the lateral lemniscus. Collaterals and probably terminals also pass from the lateral lemniscus to other nuclei in its path and receive in turn axons from these nuclei. They are the accessory nucleus, the medial preolivary nucleus, the lateral preolivary or semilunar nucleus and the nucleus of the lateral lemniscus. |
| The trapezoid body consists of horizontal fibers in the ventral part of the formatio reticularis of the lower part of the pons behind its deep transverse fibers and the pyramid bundles. The axons come from the dorsal and ventral portions of the cochlear nucleus. After crossing the raphé, where they decussate with those from the opposite side, they turn upward to form the lateral lemniscus. Fibers from the striæ medullares contribute to the trapezoid body, in addition it sends terminals or collaterals to and receives axons from the superior olivary nucleus, the nucleus of the trapezoid body, the lateral preolivary or semilunar nucleus and the mesial preolivary nucleus. |
| The cochlear nucleus, the terminal nucleus for the nerve of hearing, is usually described as consisting of a larger dorsal nucleus on the dorsal and lateral aspect of the inferior peduncle forming a prominent projection, the acoustic tubercle, and a ventral or accessory cochlear nucleus more ventral to the inferior peduncle. The two nuclei are continuous and are merely portions of one large nucleus. The axons from cells of the spiral ganglion of the cochlear nerve on reaching the nucleus divide into ascending and descending branches which enter the ventral and dorsal nuclei respectively. Axons from the large fusiform cells of the dorsal nucleus pass partly by way of the striæ medullares to the trapezoid body and lateral lemniscus and the nuclei associated with the former, and partly transversely beneath the inferior peduncle and spinal tract of the trigeminal to the trapezoid body. Axons from the ventral cochlear nucleus pass partly by the striæ medullares but for the most part horizontally to the trapezoid body. |
| The superior olivary nucleus is a small mass of gray matter situated on the dorsal surface of the lateral part of the trapezoid body. Some of its axons pass backward to the abducent nucleus, this bundle is known as the peduncle of the superior olivary nucleus. Other fibers from the nucleus join the posterior longitudinal bundle and terminate in the nuclei of the trochlear and oculomotor nerves. The majority of its axons, after giving off collaterals to the nucleus itself join the lateral lemniscus of the same side, other axons pass in the trapezoid body toward the ventral portion of the cochlear nucleus. |
| The nucleus of the trapezoid body lies between the root fibers of the abducent nerve and the superior olivary nucleus. Its cells lie among the fibers of the trapezoid body. In it terminate fibers and collaterals of the trapezoid body which come from the cochlear nucleus of the opposite and probably the same side and from the opposite trapezoid nucleus. They terminate in the nucleus of the trapezoid body in diffuse arborizations and peculiar end plaques or acoustic calyces of yellowish color which fuse with the cell bodies. Its cells are round and of medium size; their axons pass into the trapezoid body, cross the median line and probably join the lateral fillet. |
| The lateral preolivary or semilunar nucleus lies ventral to the superior olivary nucleus. In it end terminals and collaterals of the trapezoid body and probably fibers of the opposite cochlear nucleus. Its axons mingle with the trapezoid body and join the lateral fillet. |
| The mesial preolivary nucleus is in contact with the ventral side of the nucleus of the trapezoid body. It receives many collaterals from the trapezoid body. Its cells are smaller than those of the trapezoid nucleus, their axons join the lateral fillet. |
| The lateral lemniscus (lateral fillet), the continuation upward of the central path of hearing, consists of fibers which come from the cochlear nuclei of the same and the opposite side by way of the trapezoid body and from the preolivary nuclei. It lies in the ventral or ventro-lateral part of the reticular formation of the pons, at first ventral then lateral to the median fillet. Above the pons these ascending fibers come to the surface at the side of the reticular formation in the trigonum lemnisci and are covered by a layer of ependyma. This part of the lateral lemniscus is known as the fillet of Reil. On reaching the level of the inferior colliculus the dorsal fibers which overlie the superior peduncle decussate in the velum medullare anterius with similar fibers of the opposite side. Numerous small masses of cells are scattered along the path of the lateral lemniscus above the superior olivary nucleus and constitute lower and upper nuclei of the lateral lemniscus. They are supplied with many collaterals and possibly terminals from the fibers of the lemniscus. The axons of the lower nucleus of the lateral lemniscus, which arise from the larger stellate or spindle-shaped cells, with long, smooth, much branched dendrites, are said by some authors to join the lateral lemniscus, but according to Cajal they pass medially toward the raphé; their termination is unknown. The cells of the upper nucleus of the lateral lemniscus are more scattered. The same uncertainty exists in regard to their termination. |
| The fibers of the lateral lemniscus end by terminals or collaterals in the inferior colliculus and the medial geniculate body. A few of the fibers are said to pass by the inferior colliculus to terminate in the middle portion of the stratum griseum of the superior colliculus, and are probably concerned with reflex movements of the eyes depending on acoustic stimuli. |
| The inferior colliculi (lower or posterior quadrigeminal bodies) are important auditory reflex centers. Each consists of a compact nucleus of gray matter covered by a superficial white layer and separated from the central gray matter about the aqueduct by a thin, deep, white layer. Many of the axons which appear in the superficial white layer ascend through the inferior brachium to the medial geniculate body. Others mainly from large cells in the dorso-mesial part of the nucleus pass through the deep white layer into the tegmentum of the same and the opposite side and descend. Their termination is unknown, but they probably constitute an auditory reflex path to the lower motor centers, perhaps descending into the spinal cord with the tectospinal fasciculus. Other axons are said to descend in the lateral lemniscus to the various nuclei in the auditory path (Held) and probably to motor nuclei of the medulla and spinal cord. |
| The medial geniculate body receives terminals and collaterals from the lateral lemniscus (the central auditory path) and also large numbers of axons from the inferior colliculus of the same side and a few from the opposite side. It is thus a station in the central auditory path. A large proportion of its axons pass forward beneath the optic tract to join the corona radiata and then sweep backward and lateralward as the auditory radiation to terminate in the cortex of the superior temporal gyrus. V. Monakow holds that Golgi cells type II are interpolated between the terminations of the incoming fibers to the medial geniculate body and the cells located there which give rise to the fibers of the auditory radiation. the medial geniculate bodies are united by the long, slender commissure of Gudden. These fibers join the optic tract as it passes over the edge of the medial geniculate and passes through the posterior part of the optic chiasma. It is probably a commissure connected with the auditory system. |
| The Vestibular Nerve (vestibular root, VIII cranial) arise from the bipolar cells in the vestibular ganglion (Scarpa’s ganglion). The peripheral fibers end in the semicircular canals, the saccule and the utricle, the end-organs concerned with mechanism for the maintenance of bodily equilibrium. The central fibers enter the medulla oblongata and pass between the inferior peduncle and the spinal tract of the trigeminal. They bifurcate into ascending and descending branches as do the dorsal root fibers of all the spinal nerves and all afferent cranial nerves. The descending branches terminate in the dorsal (medial) vestibular nucleus, the principal nucleus of the vestibular nerve. This nucleus is prolonged downward into a descending portion in which end terminals and collaterals of the descending branch. The ascending branches pass to Deiters’s nucleus, to Bechterew’s nucleus and through the inferior peduncle of the cerebellum to the nucleus tecti of the opposite side. |
| The dorsal vestibular nucleus (medial or principal nucleus) is a large mass of small cells in the floor of the fourth ventricle under the area acustica, located partly in the medulla and partly in the pons. The striæ medullares cross the upper part of it. It is separated from the median plane by the nucleus intercalatus. Its axons pass into the posterior longitudinal bundle of the same and the opposite side and ascend to terminate in the nucleus abducens of the same side and in the trochlear nucleus and the oculo-motor nucleus of the opposite side, and to the motor nuclei of the trigeminal on both sides. The descending portion, the nucleus of the descending tract extends downward as far as the upper end of the nucleus gracilis, and the decussation of the medial lemniscus. It is sometimes called the inferior vestibular nucleus. Many of its axons cross the midline and probably ascend with the medial lemniscus to the ventro-lateral region of the thalamus. |
 |
| FIG. 761– Terminal nuclei of the vestibular nerve, with their upper connections. (Schematic.) 1. Cochlear nerve, with its two nuclei. 2. Accessory nucleus. 3. Tuberculum acusticum. 4. Vestibular nerve. 5. Internal nucleus. 6. Nucleus of Deiters. 7. Nucleus of Bechterew. 8. Inferior or descending root of acoustic. 9. Ascending cerebellar fibers. 10. Fibers going to raphé. 11. Fibers taking an oblique course. 12. Lemniscus. 13. Inferior sensory root of trigeminal. 14. Cerebrospinal fasciculus. 15. Raphé. 16. Fourth ventricle. 17. Inferior peduncle. Origin of striæ medullares. (Testut.) |
| |
| The lateral vestibular nucleus (Deiters’s nucleus) is the continuation upward and lateralward of the principal nucleus, and in it terminate many of the ascending branches of the vestibular nerve. It consists of very large multipolar cells whose axons form an important part of the posterior longitudinal bundle of the same and the opposite side. The axons bifurcate as they enter the posterior longitudinal bundle, the ascending branches send terminals and collaterals to the motor nuclei of the abducens, trochlear and oculomotor nerves, and are concerned in coördinating the movements of the eyes with alterations in the position of the head; the descending branches pass down in the posterior longitudinal bundle into the anterior funiculus of the spinal cord as the vestibulospinal fasciculus (anterior marginal bundle) and are distributed to motor nuclei of the anterior column by terminals and collaterals. Other fibers are said to pass directly to the vestibulospinal fasciculus without passing into the posterior longitudinal bundle. The fibers which pass into the vestibulospinal fasciculus are intimately concerned with equilibratory reflexes. Other axons from Deiters’s nucleus are supposed to cross and ascend in the opposite medial lemniscus to the ventro-lateral nuclei of the thalamus; still other fibers pass into the cerebellum with the inferior peduncle and are distributed to the cortex of the vermis and the roof nuclei of the cerebellum; according to Cajal they merely pass through the nucleus fastigii on their way to the cortex of the vermis and the hemisphere. |
| The superior vestibular nucleus (Bechterew’s nucleus) is the dorso-lateral part of the vestibular nucleus and receives collaterals and terminals from the ascending branches of the vestibular nerve. Its axons terminate in much the same manner as do those from the lateral nucleus. |
| The Facial Nerve (VII cranial) consists of somatic sensory, sympathetic afferent, taste, somatic motor and sympathetic efferent fibers. The afferent or sensory fibers arise from cells in the geniculate ganglion. This portion of the nerve is often described as the nervus intermedius. |
| (1) The somatic sensory fibers are few in number and convey sensory impulses from the middle ear region. Their existence has not been fully confirmed. Their central termination is likewise uncertain, it is possible that they join the spinal tract of the trigeminal as do the somatic sensory fibers of the vagus and glossopharyngeal. |
| (2) The sympathetic afferent fibers are likewise few in number and of unknown termination. |
| (3) Taste fibers convey impulses from the anterior two-thirds of the tongue via the chorda tympani. They are supposed to join the tractus solitarius and terminate in its nucleus. The central connections of this nucleus have already been considered. |
| (4) Somatic motor fibers, supplying the muscles derived from the hyoid arch, arise from the large multipolar cells of the nucleus of the facial nerve. This nucleus is serially homologous with the nucleus ambiguus and lateral part of the anterior column of the spinal cord. Voluntary impulses from the cerebral cortex are conveyed by terminals and collaterals of the pyramidal tract of the opposite side, indirectly, that is with the interpolation of a connecting neuron, to the facial nucleus. This nucleus undoubtedly receives many reflex fibers from various sources, i. e., from the superior colliculus via the ventral longitudinal bundle (tectospinal fasciculus) for optic reflexes; from the inferior colliculus via the auditory reflex path; and indirectly from the terminal sensory nuclei of the brain-stem. Through the posterior longitudinal bundle it is intimately connected with other motor nuclei of the brain-stem. |
| (5) Sympathetic efferent fibers (preganglionic fibers) arise according to some authors from the small cells of the facial nucleus, or according to others from a special nucleus of cells scattered in the reticular formation, dorso-medial to the facial nucleus. This is sometimes called the superior salivatory nucleus. These preganglionic fibers are distributed partly via the chorda tympani and lingual nerves to the submaxillary ganglion, thence by postganglionic (vasodilator) fibers to the submaxillary and sublingual glands. Some of the preganglionic fibers pass to the sphenopalatine ganglion via the great superficial petrosal nerve. |
| The Abducens Nerve (VI cranial) contains somatic motor fibers only which supply the lateral rectus muscle of the eye. The fibers arise from the nucleus of the abducens nerve and pass ventrally through the formatio reticularis of the pons to emerge in the transverse groove between the caudal edge of the pons and the pyramid. The nucleus is serially homologous with the nuclei of the trochlear and oculomotor above and with the hypoglossal and medial part of the anterior column of the spinal cord below. It is situated close to the floor of the fourth ventricle, just above the level of the striæ medullares. Voluntary impulses from the cerebral cortex are conducted by the pyramidal tract fibers (corticopontine fibers). These fibers probably terminate in relation with association neurons which control the coördinated action of all the eye muscles. This association and coördination mechanism is interposed between the terminals and collaterals of the voluntary fibers and the neurons within the nuclei of origin of the motor fibers to the eye muscles. The fibers of the posterior longitudinal bundle are supposed to play an important role in the coördination of the movements of the eyeball. Whether it is concerned only with coördinations between the vestibular apparatus and the eye or with more extensive coördinations is unknown. Many fibers of the posterior longitudinal bundle have their origin in the terminal nuclei of the vestibular nerve and from the posterior longitudinal bundle many collaterals and terminals are given off to the abducent nucleus as well as to the trochlear and oculomotor nuclei. The abducens nucleus probably receives collaterals and terminals from the ventral longitudinal bundle (tectospinal fasciculus); fibers which have their origin in the superior colliculus, the primary visual center, and are concerned with visual reflexes. Others probably come from the reflex auditory center in the inferior colliculus and from other sensory nuclei of the brain-stem. |
| The Trigeminal Nerve (V cranial) contains somatic motor and somatic sensory fibers. The motor fibers arise in the motor nucleus of the trigeminal and pass ventro-laterally through the pons to supply the muscles of mastication. The sensory fibers arise from the unipolar cells of the semilunar ganglion; the peripheral branches of the T-shaped fibers are distributed to the face and anterior two-thirds of the head; the central fibers pass into the pons with the motor root and bifurcate into ascending and descending branches which terminate in the sensory nuclei of the trigeminal. |
| The motor nucleus of the trigeminal is situated in the upper part of the pons beneath the lateral angle of the fourth ventricle. It is serially homologous with the facial nucleus and the nucleus ambiguus (motor nucleus of the vagus and glossopharyngeal) which belong to the motor nuclei of the lateral somatic group. The axons arise from large pigmented multipolar cells. The motor nucleus receives reflex collaterals and terminals, (1) from the terminal nucleus of the trigeminal of the same and a few from the opposite side, via the central sensory tract (trigeminothalamic tract); (2) from the mesencephalic root of the trigeminal; (3) from the posterior longitudinal bundle; (4) and probably from fibers in the formatio reticularis. It also receives collaterals and terminals from the opposite pyramidal tract (corticopontine fibers) for voluntary movements. There is probably a connecting or association neuron interposed between these fibers and the motor neurons. |
| The terminal sensory nucleus consists of an enlarged upper end, the main sensory nucleus, and a long more slender descending portion which passes down through the pons and medulla to become continuous with the dorsal part of the posterior column of the gray matter especially the substantia gelatinosa of the spinal cord. This descending portion consists mainly of substantia gelatinosa and is called the nucleus of the spinal tract of the trigeminal nerve. |
| The main sensory nucleus lies lateral to the motor nucleus beneath the superior peduncle. It receives the short ascending branches of the sensory root. The descending branches which form the tractus spinalis, pass down through the pons and medulla on the lateral side of the nucleus of the tractus spinalis, in which they end by collaterals and terminals, into the spinal cord on the level of the second cervical segment. It decreases rapidly in size as it descends. At first it is located between the emergent part of the facial nerve and the vestibular nerve, then between the nucleus of the facial nerve and the inferior peduncle. Lower down in the upper part of the medulla it lies beneath the inferior peduncle and is broken up into bundles by the olivocerebellar fibers and the roots of the ninth and tenth cranial nerves. Finally it comes to the surface of the medulla under the tubercle of Rolando and continues in this position lateral to the fasciculus cuneatus as far as the upper part of the cervical region where it disappears. |
| The cells of the sensory nucleus are of large and medium size and send their axons into the formatio reticularis where they form a distinct bundle, the central path of the trigeminal (trigeminothalamic tract), which passes upward through the formatio reticularis and tegmentum to the ventro-lateral part of the thalamus. Most of the fibers cross to the trigeminothalamic tract of the opposite side. This tract lies dorsal to the medial fillet; approaches close to it in the tegmentum and terminates in a distinct part of the thalamus. From the thalamus impulses are conveyed to the somatic sensory area of the cortex by axons of cells in the thalamus through the internal capsule and corona radiata. Many collaterals are given off in the medulla and pass from the trigeminothalamic tract to the motor nuclei, especially to the nucleus ambiguus, the facial nucleus and the motor nucleus of the trigeminal. |
| The somatic sensory fibers of the vagus, the glossopharyngeal and the facial nerves probably end in the nucleus of the descending tract of the trigeminal and their cortical impulses are probably carried up in the central sensory path of the trigeminal. |
| The mesencephalic root (descending root of the trigeminal) arises from unipolar cells arranged in scattered groups in a column at the lateral edge of the central gray matter surrounding the upper end of the fourth ventricle and the cerebral aqueduct. They have usually been considered as motor fibers that join the motor root, but Johnston claims that they join the sensory root of the trigeminal, that they develop in the alar, not in the basal lamina, and that the pear-shaped unipolar cells are sensory in type. |
| The Trochlear Nerve (IV cranial) contains somatic motor fibers only. It supplies the superior oblique muscle of the eye. Its nucleus of origin, trochlear nucleus, is a small, oval mass situated in the ventral part of the central gray matter of the cerebral aqueduct at the level of the upper part of the inferior colliculus. The axons from the nucleus pass downward in the tegmentum toward the pons, but turn abruptly dorsalward before reaching it, and pass into the superior medullary velum, in which they cross horizontally, to decussate with the nerve of the opposite side, and emerges from the surface of the velum, immediately behind the inferior colliculus. The cells of the trochlear nucleus are large, irregular and yellowish in color. The nuclei of the two sides are separated by the raphé through which dendrites extend from one nucleus to the other. They receive many collaterals and terminals from the posterior longitudinal bundle which lies on the ventral side of the nucleus. |
| There are no branches from the fibers of the pyramidal tracts to these nuclei; the volitional pathway must be an indirect one, as is the case with other motor nuclei. |
| The Oculomotor Nerve (III cranial) contains somatic motor fibers to the Obliquus inferior, Rectus inferior, Rectus superior, Levator palpebræ superioris and Rectus medialis muscles and sympathetic efferent fibers (preganglionic fibers) to the ciliary ganglion. The postganglionic fibers connected with these supply the ciliary muscle and the sphincter of the iris. The axons arise from the nucleus of the oculomotor nerve and pass in bundles through the posterior longitudinal bundle, the tegmentum, the red nucleus and the medial margin of the substantia nigra in a series of curves and finally emerge from the oculomotor sulcus on the medial side of the cerebral peduncle. |
| The oculomotor nucleus lies in the gray substance of the floor of the cerebral aqueduct subjacent to the superior colliculus and extends in front of the aqueduct a short distance into the floor of the third ventricle. The inferior end is continuous with the trochlear nucleus. It is from 6 to 10 mm. in length. It is intimately related to the posterior longitudinal bundle which lies against its ventro-lateral aspect and many of its cells lie among the fibers of the posterior longitudinal bundle. The nucleus of the oculomotor nerve contains several distinct groups of cells which differ in size and appearance from each other and are supposed to send their axons each to a separate muscle. Much uncertainty still exists as to which group supplies which muscle. There are seven of these groups or nuclei on either side of the midline and one medial nucleus. The cells of the anterior nuclei are smaller and are supposed to give off the sympathetic efferent axons. The majority of fibers arise from the nucleus of the same side some, however, cross from the opposite side and are supposed to supply the Rectus medialis muscle. Since oculomotor and abducens nuclei are intimately connected by the posterior longitudinal bundle this decussation of fibers to the Medial rectus may facilitate the conjugate movements of the eyes in which the Medial and Lateral recti are especially involved. |
| Many collaterals and terminals are given off to the oculomotor nucleus from the posterior longitudinal bundle and thus connect it with the vestibular nucleus, the trochlear and abducens nuclei and probably with other cranial nuclei. Fibers from the visual reflex center in the superior colliculus pass to the nucleus. It is also connected with the cortex of the occipital lobe of the cerebrum by fibers which pass through the optic radiation. The pathway for voluntary motor impulses is probably similar to that for the abducent nerve. |
| The Optic Nerve or Nerve of Sight (II cranial) consists chiefly of coarse fibers which arise from the ganglionic layer of the retina. They constitute the third neuron in the series composing the visual path and are supposed to convey only visual impressions. A number of fine fibers also pass in the optic nerve from the retina to the primary centers and are supposed to be concerned in the pupillary reflexes. There are in addition a few fibers which pass from the brain to the retina; they are supposed to control chemical changes in the retina and the movements of the pigment cells and cones. Each optic nerve has, according to Salzer, about 500,000 fibers. |
 |
| FIG. 762– Figure showing the different groups of cells, which constitute, according to Perlia, the nucleus of origin of the oculomotor nerve. 1. Posterior dorsal nucleus. 1’. Posterior ventral nucleus. 2. Anterior dorsal nucleus. 2’. Anterior ventral nucleus. 3. Central nucleus. 4. Nucleus of Edinger and Westphal. 5. Antero-internal nucleus. 6. Antero-external nucleus. 8. Crossed fibers. 9. Trochlear nerve, with 9’, its nucleus of origin, and 9", its decussation. 10. Third ventricle. M, M. Median line. (Testut.) |
| |
| In the optic chiasma the nerves from the medial half of each retina cross to enter the opposite optic tract, while the nerves from the lateral half of each retina pass into the optic tract of the same side. The crossed fibers tend to occupy the medial side of each optic nerve, but in the chiasma and in the optic tract they are more intermingled. The optic tract is attached to the tuber cinereum and lamina terminalis and also to the cerebral peduncle as it crosses obliquely over its under surface. These are not functional connections. A small band of fibers from the medial geniculate body joins the optic tract as the latter passes over it and crosses to the opposite tract and medial geniculate body in the posterior part of the chiasma. This is the commissure of Gudden and is probably connected with the auditory system. |
| Most of the fibers of the optic tract terminate in the lateral geniculate body, some pass through the superior brachium to the superior colliculus, and others either pass over or through the lateral geniculate body to the pulvinar of the thalamus. These end-stations are often called the primary visual centers. |
| The lateral geniculate body consists of medium-sized pigmented nerve cells arranged in several layers by the penetrating fibers of the optic tract. Their axons pass upward beneath the longer fibers of the optic tract, the tænia semicircularis, the caudate nucleus and the posterior horn of the lateral ventricle where they join the optic radiation of Gratiolet. They pass backward and medially to terminate in the visuo-sensory cortex in the immediate neighborhood of the calcarine fissure of the occipital lobe. This center is connected with the one in the opposite side by commissural fibers which course in the optic radiation and the splenium of the corpus callosum. Association fibers connect it with other regions of the cortex of the same side. |
 |
| FIG. 763– Scheme showing central connections of the optic nerves and optic tracts. |
| |
| The region of the pulvinar in which optic tract fibers terminate resembles in structure the lateral geniculate body. Its axons also have a similar course though in a somewhat more dorsal plane. |
| The superior colliculus receives fibers from the optic tract through the superior brachium. Some enter by the superficial white layer (stratum zonale), others appear to dip down into the gray cap (stratum cinereum) while others probably decussate across the midline to the opposite colliculus. Other fibers from the superior brachium pass into the stratum opticum (upper gray-white layer). Some of these turn upward into the gray cap while others terminate among the cells of this layer. Since the superior colliculi appear to be the central organs concerned in the control of eye-muscle movements and eye-muscle reflexes we should expect to find them receiving fibers from other sensory paths. Many fibers pass to the superior colliculus from the medial fillet as the latter passes through the tegmentum bringing the superior colliculus into relation with the sensory fibers of the spinal cord. Fibers from the central sensory path of the trigeminal probably pass with these. Part of the ventral spinocerebellar tract (Gowers) is said to pass up through the reticular formation of the pons and mid-brain toward the superior colliculus and the thalamus. The superior colliculus is intimately connected with the central auditory path (the lateral lemniscus), as part of its fibers pass the inferior colliculus and terminate in the superior colliculus. They are probably concerned with reflex movements of the eyes depending on auditory stimuli. The superior colliculus is said to receive fibers from the stria medullaris thalamis of the opposite side which pass through the commissura habenulæ and turn back to the roof of the mid-brain, especially to the superior colliculus. By this path both the primary and cortical olfactory centers are brought into relation with the eye-muscle reflex apparatus. |
| The fibers which pass to the nuclei of the eye muscles arise from large cells in the stratum opticum and stratum lemnisci and pass around the ventral aspect of the central gray matter where most of them cross the midline in the fountain decussation of Meynert, and then turn downward to form the ventral longitudinal bundle. This bundle runs down partly through the red nucleus, in the formatio reticularis, ventral to the posterior longitudinal bundle of the mid-brain, pons and medulla oblongata into the ventral funiculus of the spinal cord where it is known as the tectospinal fasciculus. Some of the fibers are said to pass down with the rubrospinal tract in the lateral funiculus. Some fibers do not decussate but pass down in the ventral longitudinal bundle of the same side on which they arise unless possibly they come from the opposite colliculus over the aqueduct. From the ventral longitudinal bundle collaterals are given off to the nuclei of the eye muscles, the oculomotor, the trochlear and the abducens. Many collaterals pass to the red nucleus, and are probably concerned with the reflexes of the rubrospinal tract. The fibers of the tectospinal tract end by collaterals and terminals either directly or indirectly among the motor cells in the anterior column of the spinal cord. |
| The superior colliculus receives fibers from the visual sensory area of the occipital cortex; they pass in the optic radiation. Probably no fibers pass from the superior colliculus to the visual sensory cortex. |
| The Olfactory Nerves (I cranial) or nerves of smell arise from spindle-shaped bipolar cells in the surface epithelium of the olfactory region of the nasal cavity. The non-medullated axons pass upward in groups through numerous foramina in the cribriform plate to the olfactory bulb; here several fibers, each ending in a tuft of terminal filaments, come into relation with the brush-like end of a single dendrite from a mitral cell. This interlacing gives rise to the olfactory glomeruli of the bulb. The termination of several or many olfactory fibers in a single glomerulus where they form synapses with the dendrites of one or two mitral cells provides for the summation of stimuli in the mitral cells and accounts in part at least for the detection by the olfactory organs of very dilute solutions. Lateral arborizations of the dendrites of the mitral cells and the connection of neighboring glomeruli by the axons of small cells of the glomeruli and the return of impulses of the mitral cells by collaterals either directly or through the interpolation of granule cells to the dendrites of the mitral cells reinforce the discharge of the mitral cells along their axons. The axons turn abruptly backward in the deep fiber layer of the bulb to form the olfactory tract. The olfactory tract is continued into the olfactory trigone, just in front of the anterior perforated substance. The axons of the mitral cells on reaching the olfactory trigone separate into three bundles, the lateral olfactory stria, the medial olfactory stria and the less marked intermedial olfactory stria. |
| The lateral olfactory striæ curve lateralward, a few of the fibers end in the olfactory trigone and the antero-lateral portion of the anterior perforated substance. Most of the fibers, however, pass into the uncus, the anterior end of the hippocampal gyrus, and there end in the complicated cortex of the hippocampal gyri. The lateral striæ more or less disappear as they cross the antero-lateral region of the anterior perforated substance. |
| The greater mass of the fibers of the olfactory tract pass into the lateral stria. Numerous collaterals are given into the plexiform layer of the subfrontal cortex, over which the striæ pass on their way to the uncus, where they intermingle with the apical dendrons of the medium-sized and small pyramidal cells of the pyramidal layer of this subfrontal or frontal olfactory cortex. The axons give rise to projection fibers which take an antero-posterior direction to the subthalamic region sending collaterals and terminal branches to the stria medullaris and others toward the thalamus. Some of the fibers extend farther back and are believed to reach the pons and medulla oblongata. |
| Most of the fibers of the lateral olfactory stria pass to the hippocampal region of the cortex, especially to the gyrus hippocampi, which may be regarded as the main ending place of the secondary olfactory path derived from axons of the mitral cells. |
| The fibers of the medial olfactory striæ terminate for the most part in the parolfactory area (Broca’s area), a few end in the subcallosal gyrus and a few in the anterior perforated substance and the adjoining part of the septum pellucidum. Some of the fibers pass into the anterior commissure (pars olfactoria) to the olfactory tract of the opposite side where they end partly within the granular layer and partly in the neighborhood of the glomeruli of the olfactory bulb, thus connecting the bulbs of the two sides. |
| The intermediate olfactory striæ are as a rule scarcely visible, the fibers terminate in the anterior perforated substance, a few are said to continue to the uncus. |
| The trigonum olfactorium, anterior perforated substance and the adjoining part of the septum pellucidum are important primary olfactory centers, especially for olfactory reflexes; in these centers terminate many axons from the mitral cells of the olfactory bulb. In addition the gray substance of the olfactory tract and the gyrus subcallosus receive terminals of the mitral cells. |
| The pathways from these centers to lower centers in the brain-stem and spinal cord are only partially known. The most direct path, the tractus olfactomesencephalicus (basal olfactory bundle of Wallenburg), is supposed to arise from cells in the gray substance of the olfactory tract, the olfactory trigone, the anterior perforated substance and the adjoining part of the septum pellucidum. The fibers are said to pass direct to the tuber cinereum, to the corpus mammillare, to the brainstem and the spinal cord. The fibers which enter the mammillary body probably come into relation with cells whose axons give rise to the fasciculus mammillo-tegmentalis (mammillo-tegmental bundle of Gudden) which is supposed to end in the gray substance of the tegmentum and of the aqueduct; some of its fibers are said to join the posterior longitudinal bundle and others to extend as far as the reticular formation of the pons. |
| Some of the fibers of the medial olfactory stria came into relation with cells in the parolfactory area of Broca and in the anterior perforated substance, whose axons course in the medullary stria of the thalamus. As the axons pass through the lower part of the septum pellucidum they are joined by other fibers whose cells receive impulses from the mitral cells. These fibers of the medullary stria end for the most part in the habenular nucleus of the same side, some, however, cross in the habenular commissure (dorsal part of the posterior commissure) to the habenular nucleus of the opposite side. A few fibers of the medullary stria are said to pass by the habenular nucleus to the roof of the mid-brain, especially the superior colliculus, while a few others come into relation with the posterior longitudinal bundle and association tracts of the mesencephalon. |
| The ganglion of the habenulæ located in the trigonum habenulæ just in front of the superior colliculus contains a mesial nucleus with small cells and a lateral nucleus with larger cells. The axons of these cells are grouped together in a bundle, the fasciculus retroflexus of Meynert, which passes ventrally medial to the red nucleus and terminates in a small medial ganglion in the substantia perforata posterior, immediately in front of the pons, called the interpeduncular ganglion. |
| The interpeduncular ganglion has rather large nerve cells whose axons curve backward and downward as the tegmental bundle of Gudden, to end partly in the dorsal tegmental nucleus and surrounding gray substance where they come into relation with association neurons and the dorsal longitudinal bundle of Schütz. |
| The majority of the axons that arise from the mitral cells of the olfactory bulb and course in the olfactory tract course in the lateral olfactory stria to the uncus and hippocampal gyrus, and terminate in the cortex. Other fibers probably pass to the uncus and hippocampal gyrus from the primary olfactory centers in the trigonum and anterior perforated substance. The gyrus hippocampus is continued through the isthmus into the gyrus cinguli which passes over the corpus callosum to the area parolfactoria. The cortical portions of these gyri are connected together by a thick association bundle, the cingulum, that lies buried in the depth of the gyrus cinguli extending forward to the parolfactory area and backward into the hippocampal region. The axons from the gyrus cinguli pass into the cingulum, many of them bifurcate, the anterior branches together with the axons which run in that direction are traceable as far forward as the anterior part of the septum pellucidum and the anterior end of the corpus striatum, where some of them are incorporated with projection fibers passing toward the internal capsule. The branches and axons which pass backward terminate partly in the hippocampus, the dentate gyrus and hippocampal gyrus. Shorter association fibers connect various sections of the gyrus fornicatus (cingulate gyrus, isthmus, and hippocampal gyrus) and these with other regions of the cortex. These gyri constitute the cortical center for smell. |
| The dentate gyrus which may be considered as a modified part of the hippocampus is partially separated from the gyrus hippocampus by the hippocampal fissure and from the fimbria by the fimbrio-dentate sulcus; it is intimately connected with the hippocampal gyrus and the hippocampus. When followed backward the dentate gyrus separates from the fimbria at the splenium, loses its incisions and knobs, and as the fasciola cinerea passes over the splenium onto the dorsal surface of the corpus callosum and spreads out into a thin layer of gray substance known as the indusium, which can be traced forward around the genu of the corpus callosum into the gyrus subcallosus. The white matter of the indusium known as the medial longitudinal striæ (nerves of Lancisi) and the lateral longitudinal striæ, are related to the indusium somewhat as the cingulum is to the gyrus cinguli. Axons from the indusium pass into the longitudinal striæ, some running forward and others backward while some after entering the medial longitudinal stria, pierce the corpus callosum to join the fornix. Some of the fibers which pass forward extend around the front of the corpus callosum and the anterior commissure, then curve downward, according to Cajal, to enter the corpus striatum where they join the olfactory projection-path. Other fibers are said to arise in the parolfactory area, the gyrus subcallosus and the anterior perforated substance (diagonal band of Broca) and course backward in the longitudinal striæ to the dentate gyrus and the hippocampal region. The indusium is usually considered as a rudimentary part of the rhinencephalon. |
| The olfactory projection fibers which arise from the pyramid cells of the uncus and hippocampus and from the polymorphic cells of the dentate gyrus form a dense stratum on the ventricular surface, especially on the hippocampus, called the alveus. These fibers pass over into the fimbria and are continued into the fornix. About one-fourth of all the fibers of the fimbria are large projection fibers, the other three-fourths consist of fine commissural fibers which pass from the hippocampus of one side through the fimbria and hippocampal commissure (ventral psalterium or lyre), to the fimbria and hippocampus of the opposite side where they penetrate the pyramidal layer and terminate in the stratum radiatum. The fibers which course in the fornix pass forward and downward into the corpora mammillare where numerous collaterals are given off and a few terminate. Most of the fibers in the fornix, however, pass through the corpora, cross the middle line and turn downward in the reticular formation in which they are said to be traceable as far as the pons and possibly farther. As the fornix passes beneath the corpus callosum it receives fibers from the longitudinal striæ of the indusium and from the cingulum; these are the perforating fibers of the fornix which pass through the corpus callosum and course in the fornix toward the mammillary body. As the fornix passes the anterior end of the thalamus a few fibers are given off to the stria medullaris of the thalamus and turn back in the stria to the habenular ganglion of the same and the opposite side, having probably the same relation that the reflex fibers have which arise from the primary centers and course in the stria medullaries of the thalamus. Aside from the fibers of the fornix which pass through the mammillary body to decussate and descend (as the mammillo-mesencephalic fasciculus), many fibers are said to pass into the bundle of Vicq d’Azyr, and one bundle of fibers is said to pass from the fornix to the tuber cinereum. |
| The mammillary bodies receive collaterals and terminals then from the cortical centers via the fornix and probably other collaterals and terminals are received directly from the primary centers through the tractus olfactomesencephalicus. According to Cajal fibers also reach the mammillary body through the peduncle of the corpus mammillare from the arcuate fibers of the tegmentum and from the main fillet. The fornix probably brings the cortical centers into relation with the reflex path that runs from the primary centers to the mammillary body and the tuber cinereum. |
| The bundle of Vicq d’Azyr (mammillo-thalamic fasciculus) arises from cells in both the medial and lateral nuclei of the mammillary body and by fibers that are directly continued from the fornix. There axons divide within the gray matter; the coarser branches pass into the anterior nucleus of the thalamus as the bundle of Vicq d’Azyr, the finer branches pass downward as the mammillo-tegmental bundle of Gudden. The bundle of Vicq d’Azyr spreads out fan-like as it terminates in the anterior or dorsal nucleus of the thalamus. A few of the fibers pass through the dorsal nucleus to the angular nucleus of the thalamus. The axons from these nuclei are supposed to form part of the thalamocortical system. |
| The mammillo-tegmental bundle has already been considered under the olfactory reflex paths. |
| The amygdaloid nucleus and the tænia semicircularis (stria terminalis) probably belong to the central olfactory apparatus. The tænia semicircularis extends from the region of the anterior perforated substance to the nucleus amygdalæ. Its anterior connections are not clearly understood. Fibers are said to arise from cells in the anterior perforated substance; some of the fibers pass in front of the anterior commissure, others join the fornix for a short distance as they pass behind the anterior commissure. The two strands ultimately join to form the tænia and pass backward in the groove between the caudate nucleus and the thalamus to the amygdaloid nucleus. Other fibers are said to pass in the opposite direction from the amygdaloid nucleus to the thalamus. |
